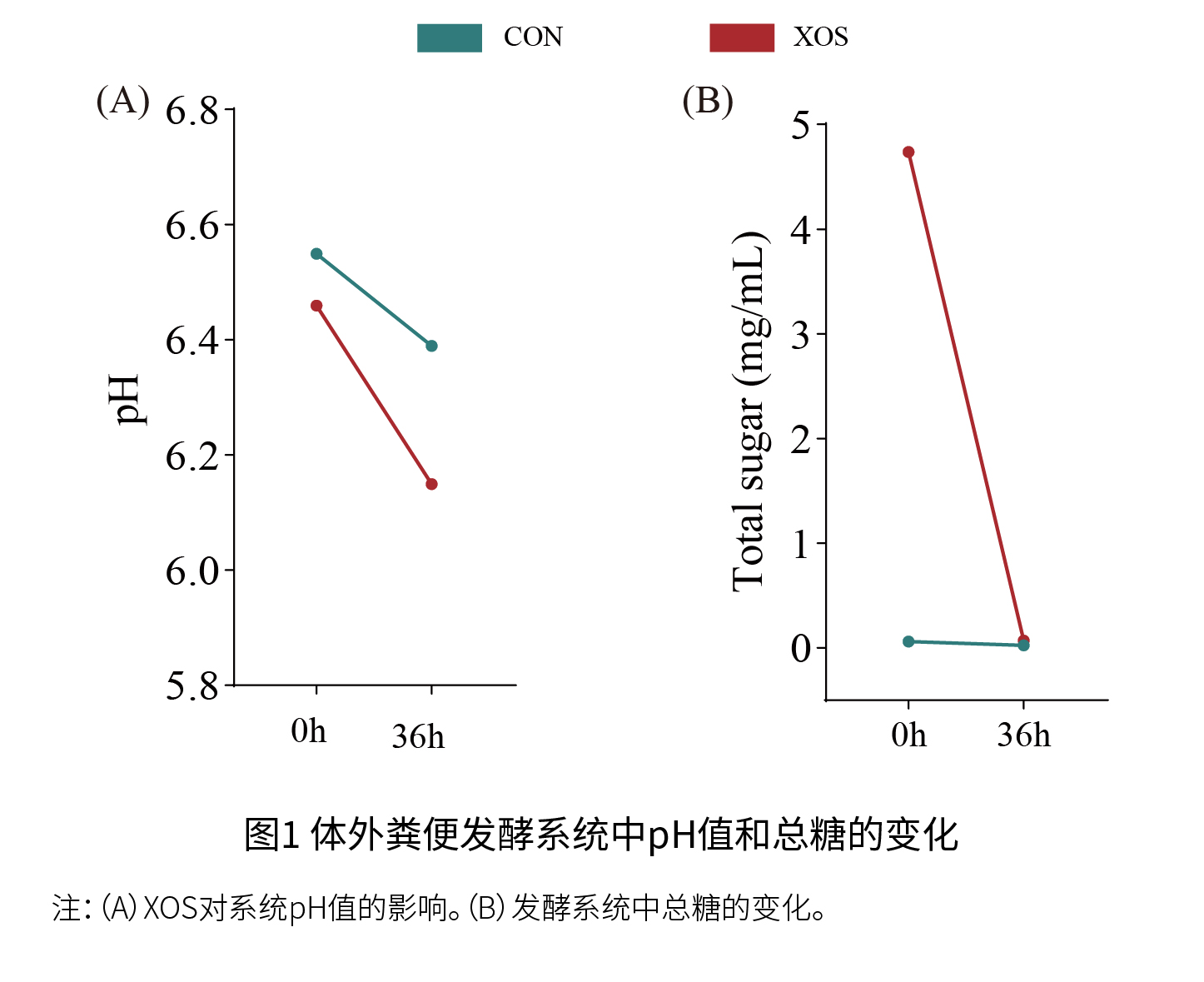
试验设计 实验材料:纯度为95%的益常青®低聚木糖。 实验一:体外粪便发酵实验 粪浆的制备:人类粪便样本来自六名健康志愿者(三名女性和三名男性),志愿者年龄在20至25岁之间,在粪便收集前至少3个月未服用任何抗生素,至少3个月未摄入任何益生元或益生菌产品,并且身体健康。在实验之前,从六名志愿者收集的粪便被转移到厌氧室进行后续处理,称重并分批混合粪便,以减少个体间差异。混合的粪便样本以1:10(w/v)的比例均匀分散在预先准备的无菌磷酸盐缓冲盐水(PBS)中,通过四层纱布过滤,所得滤液用作接种物,粪便样本的操作保持在厌氧条件下。 体外粪便发酵:XOS的体外粪便发酵在严格的厌氧条件下进行,以模拟肠道中XOS的发酵。没有添加额外糖的培养物用作对照组(CON),而添加XOS的培养物用作实验组(XOS),实验进行了五次重复。使用葡萄糖作为标准品通过酚-硫酸法评估发酵前后总糖含量,用pH计检测发酵过程中0 h和36 h的pH值,并在发酵结束时(36 h)收集粪便发酵样品进行16S rDNA测序和代谢组学分析。 16S rDNA分析:每组设置五个重复。 非靶向代谢组学分析:每组设置五个重复。 目标有机酸分析:通过超高效液相色谱-质谱联用(UPLC-MS)对粪便发酵上清液中的目标有机酸进行分析,每组设置三个重复。 短链脂肪酸(SCFAs)的测定:使用气相色谱-质谱联用(GCMS-QP2010,Shimadzu,Japan)定量SCFAs,每组设置三个重复。 色氨酸和胆汁酸分析:粪便发酵上清液中的靶向色氨酸和胆汁酸代谢物分析通过UPLC-MS进行,每组设置五个重复。 碳水化合物活性酶分析:从美国国家生物技术信息中心(NCBI)获得主要富集物种的基因组数据,下载最新的碳水化合物活化酶相关蛋白序列(CAZyDB.07262023),并构建一个本地蛋白质序列数据库。 微生物-微生物和微生物-代谢物相互作用网络分析:基于斯皮尔曼相关性分析构建。 实验二:动物实验 选择30只8周龄雄性C57BL/6J小鼠,饲养温度控制在24±1 °C,并维持12 h光照-黑暗周期,湿度维持在55±10%。 1)动物实验方案1:小鼠随机分为对照组(CON-I)和治疗组(XOS-I:600 mg/kg BW/d XOS)(每组n=5),小鼠可以自由获取标准饲料和饮用水。经过1周的适应期后,XOS-I组的小鼠通过灌胃给予600 mg/kg BW/d XOS,剂量是根据美国食品药品监督管理局(FDA)的指导原则(基于体表面积缩放)选择的,即将XOS的人体最大摄入剂量(3g /天)转换为小鼠等效剂量(~600 mg/kg),持续14天,CON-I组的小鼠通过灌胃给予等量的0.9%生理盐水。每组小鼠的粪便样本(每组样本量n=5)被收集用于16S rDNA分析和非靶向代谢组分析。 2)动物实验方案2:小鼠随机分为四组,对照组(Control)、衰老组(Model)、治疗组(XOS:600 mg/kg BW/d,衰老+XOS)和阳性对照组(精氨酸:0.4 mg/g BW/d精氨酸,衰老+精氨酸)(每组n=5),在饲喂期间小鼠可以正常饮食并自由饮水。小鼠适应1周后,对照组皮下注射0.9%生理盐水,除了对照组之外的所有组每日皮下注射1000 mg/kg BW的D-半乳糖(D-gal)以建立衰老模型,并每日通过灌胃给予0.2 mL的0.9%生理盐水、600 mg/kg BW XOS或0.4 mg/g BW/d精氨酸,共进行7周,每周记录小鼠的体重。实验结束时,每组小鼠的粪便样本(每组样本量n=5)被收集用于16S rDNA分析和非靶向代谢组分析,小鼠用异氟烷麻醉并通过颈椎脱位处死。收集大脑和0.5 cm的原发结肠组织并浸入4%甲醛(PFA)溶液中进行组织病理学分析,剩余的大脑、肝脏、血清和结肠样本储存在-80 °C。 行为测试:进行开放场测试和Morris水迷宫(MWM)测试,测试前,小鼠至少提前1 h转移到配备必要设备的房间。 组织学分析:结肠和大脑样本固定在4% PFA中并用石蜡包埋。结肠样本用苏木精-伊红(H&E)染色,大脑组织用抗离子化钙结合适配蛋白1(Iba1)和胶质纤维酸性蛋白(GFAP)染色。使用BX-41显微镜(Nikon Corporation,Japan)获取图像。 生化分析:收集大脑和肝脏组织制成10%组织匀浆,用于测定抗氧化指标。收集血清和结肠组织用于测定炎症细胞因子,根据检测试剂盒的说明(南京复星生物科技有限公司,中国),检测血清和结肠组织中的IL-1β和TNF-α水平。 试验结果 实验一:体外粪便发酵实验 (1)XOS补充后的pH和总糖变化 如图1A所示,与CON组相比,XOS组的pH下降更为明显(XOS:从0 h到36 h,CON:从0 h到36 h),表明添加XOS的发酵系统产生了酸性底物。图1B显示,XOS组的总糖含量从4.7±0.077 mg/mL(0 h)显著下降到0.071±0.0038 mg/mL(36 h),表明在发酵过程中糖被微生物完全降解。
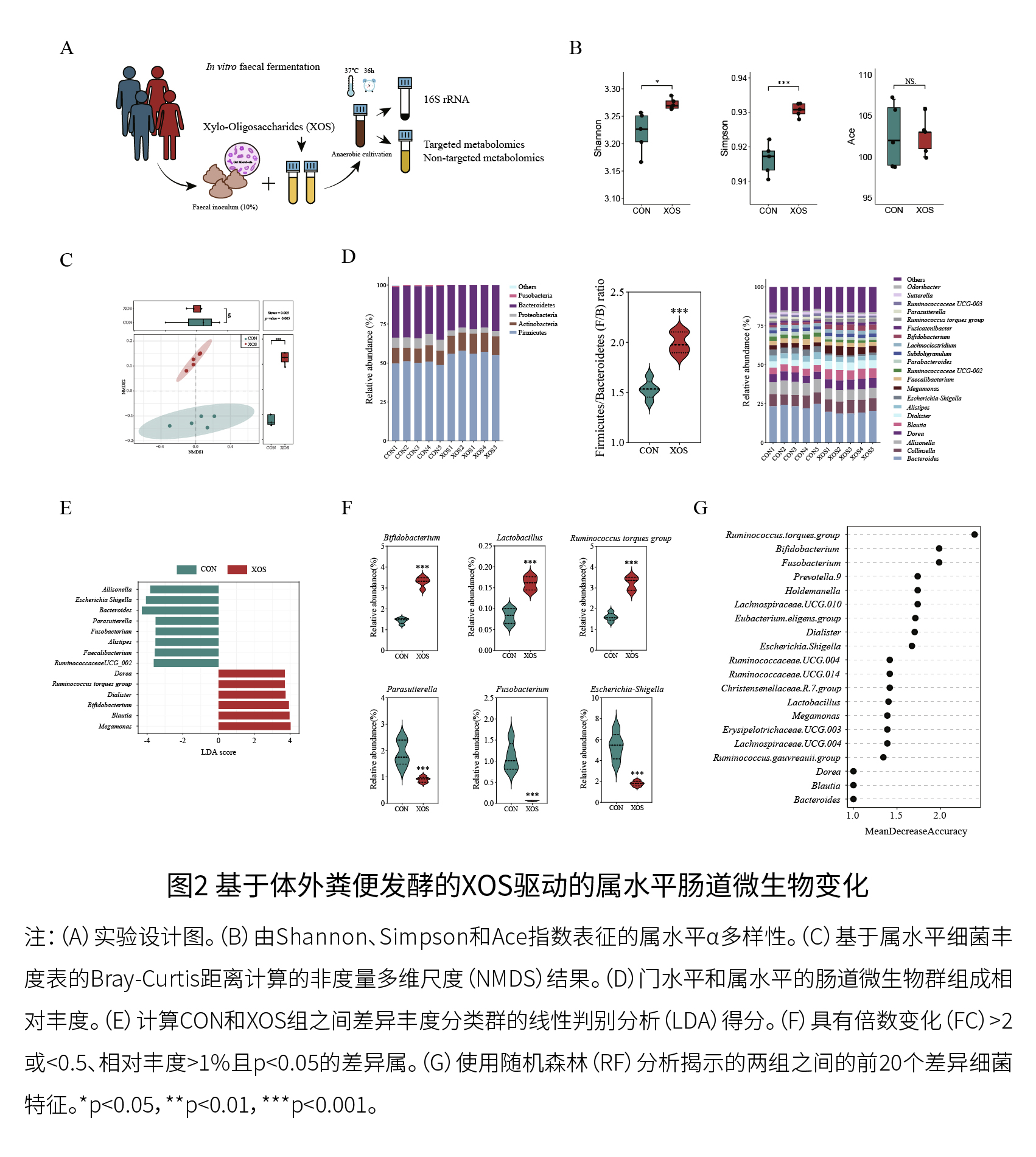
(2)鉴定对XOS有响应的关键微生物属/种 如图2A和2B所示,XOS处理使Shannon和Simpson指数显著增加(p<0.05),而Ace指数没有显著变化。图2C显示,XOS处理显著改变了微生物群组成(p=0.005)。图2D显示,XOS处理增加了厚壁菌门/拟杆菌门的比例并降低了变形菌门的相对丰度。如图2E-2G所示,CON和XOS组之间共同鉴定出的关键差异类群包括:扭链瘤胃球菌、双歧杆菌、埃希氏-志贺氏菌、梭杆菌,其中前两者在XOS组中富集,而后两者则在XOS组中减少。LEfSe分析表明,布劳特氏菌属、Dorea、戴阿利斯特杆菌属和巨单胞菌属的相对丰度在XOS处理后升高(LDA>3.5,p<0.05),乳酸杆菌在XOS组中的相对丰度也增加。
如图3C所示,在OUT水平上,24个OTUs在XOS组中富集并被鉴定到种水平,包括戴阿利斯特杆菌属(OTU003)、双歧杆菌(OTU070、OTU032)、巨单胞菌属(OTU044)、乳酸杆菌(OTU037、OTU001)等。图3D显示,在LEfSe分析中,筛选出了6个差异OTUs,其中,长双歧杆菌长亚种(OTU032)被分类到种水平,并在XOS组中富集。 总之,只有7个差异OTUs在XOS组中富集,包括动物双歧杆菌(OTU070)、长双歧杆菌长亚种(OTU032)、唾液乳杆菌(OTU001)、戈氏副拟杆菌(OTU123)、脆弱拟杆菌(OTU124)、普拉梭菌(OTU154)、发酵乳杆菌(OTU037)。
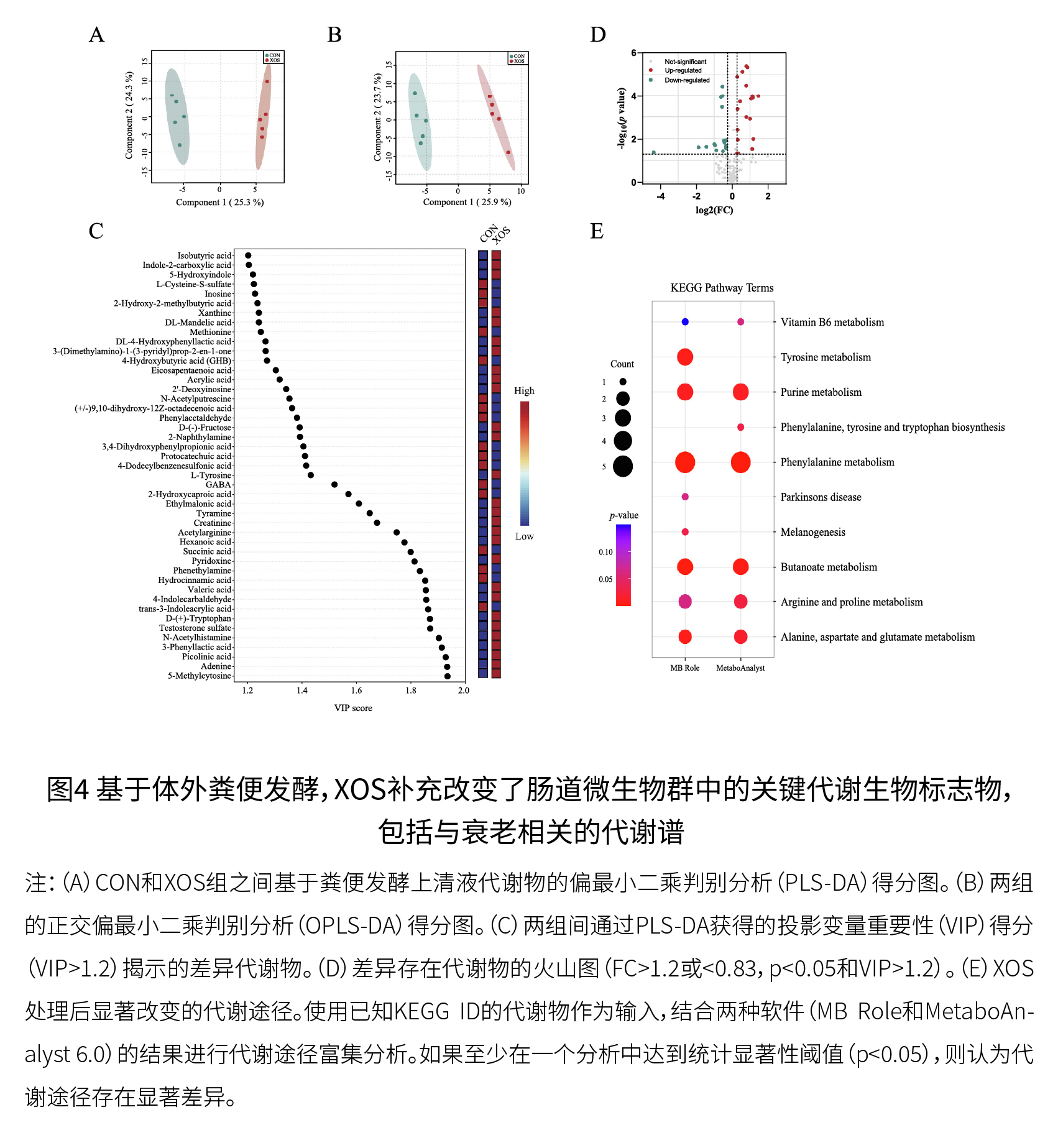
(3)鉴定出的主要XOS响应微生物都配备了利用XOS的酶 如图3F所示,所有分析的7个主要富集物种的菌株至少包含一个编码XOS降解相关酶的GH家族(GH1家族存在于所有7个主要富集物种中)。戈氏副拟杆菌(OTU123)、长双歧杆菌长亚种(OTU032)和脆弱拟杆菌(OTU124)的XOS降解GH家族的数量和/或多样性高于发酵乳杆菌(OTU037)和唾液乳杆菌(OTU001)。随后,将基因组分析的结果和这7个物种相对丰度的变化结合起来,进一步评估它们对XOS的响应,其中,长双歧杆菌长亚种(OTU032)可能是响应最强烈的物种。 (4)XOS改变了生物学上重要途径中的关键代谢生物标志物,包括与宿主衰老相关的代谢物/途径 图4D显示了XOS和对照组之间的差异代谢物(FC>1.2或<0.83,p<0.05和VIP>1.2),包括先前报道的与衰老相关的微生物产物(包括肌苷、γ-氨基丁酸和N-乙酰组胺)。如图4E所示,XOS干预影响了肠道微生物群的关键代谢谱,包括一些与衰老相关的代谢途径,如精氨酸和脯氨酸代谢、丁酸代谢和色氨酸代谢。
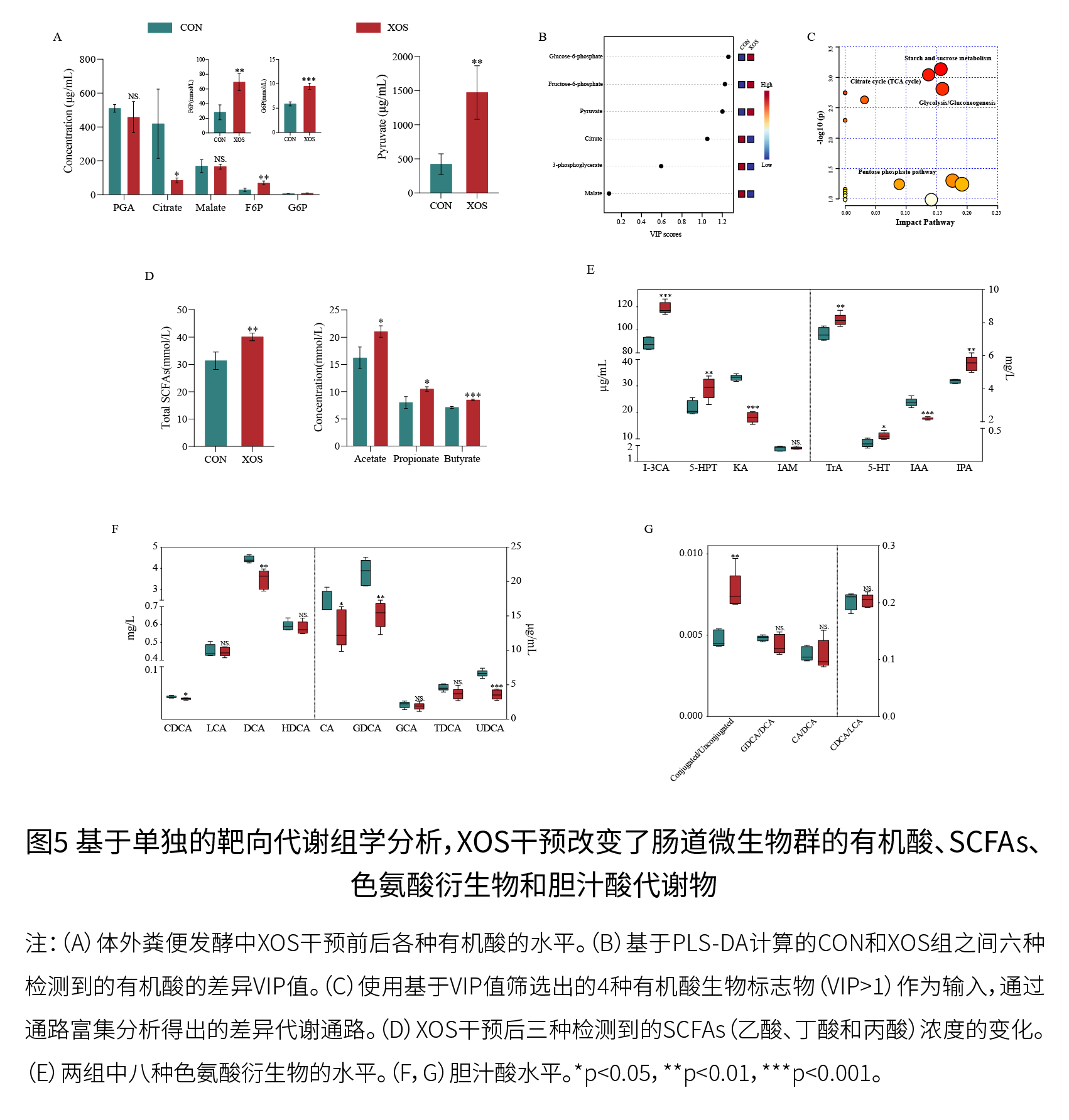
(5)XOS补充后有机酸和SCFAs的变化 如图5A所示,与CON组相比,XOS干预并未引起苹果酸和PGA水平的显著变化,柠檬酸水平显著下降,而其余有机酸的含量显著增加,G6P的含量增幅最为显著(p<0.001)。如图5B和5C所示,XOS干预调节了淀粉和蔗糖代谢、戊糖磷酸途径(PPP)和糖酵解/糖异生等代谢途径。如图5D所示,与CON组相比,XOS组中总SCFAs(乙酸、丙酸和丁酸之和)的含量显著增加,乙酸、丙酸和丁酸的浓度显著增加,其中丁酸的浓度增加最为显著(p<0.001)。XOS组和CON组中乙酸的浓度都很高。
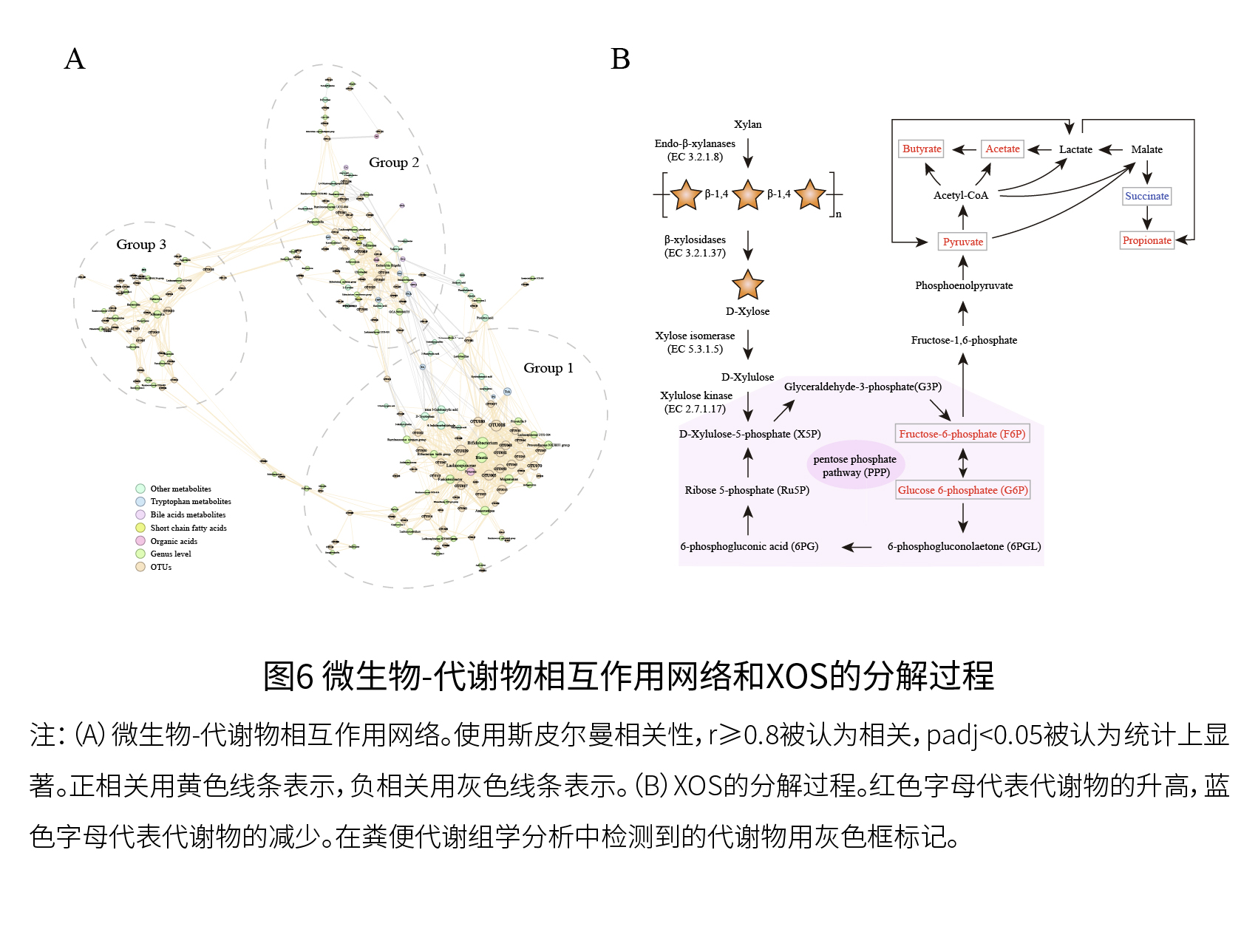
(6)色氨酸和胆汁酸代谢也因XOS而发生了显著变化 如图5E所示,与CON组相比,XOS组中色氨酸衍生物的水平增加,如吲哚-3-甲醛(I-3CA)、IPA、5-羟色氨酸(5-HPT)、色胺(TrA)和5-HT,而吲哚-3-乙酸(IAA)和犬尿氨酸(KA)的含量减少。如图5F和5G所示,XOS处理降低了脱氧胆酸(DCA)、胆酸(CA)、甘氨脱氧胆酸(GDCA)、石胆酸(LCA)、熊去氧胆酸(UDCA)和鹅去氧胆酸(CDCA)的含量,而结合型/非结合型胆汁酸的比值则有所增加。与CON组相比,XOS处理在CDCA/LCA和CA/DCA比例上没有显著差异,这表明添加XOS可能对胆汁酸的去羟基化影响较小。色氨酸代谢和胆汁酸代谢是调节人类健康和衰老的重要代谢途径,XOS处理显著影响这些代谢物的丰度。 (7)构建了微生物-代谢物相互作用网络,并提出了XOS的分解代谢途径 如图6B所示,XOS通过双歧杆菌、布劳特氏菌和Anaerostipes等细菌利用β-木糖苷酶(EC 3.2.1.37)分解为D-木糖,具有木糖异构酶(EC 5.3.1.5)和木酮糖激酶(EC 2.7.1.17)等酶的肠道微生物将D-木糖代谢为D-木酮糖-5-磷酸,进入PPP途径,并有助于F6P和G6P的产生,刺激丙酮酸的产生,最终导致SCFAs水平的增加。
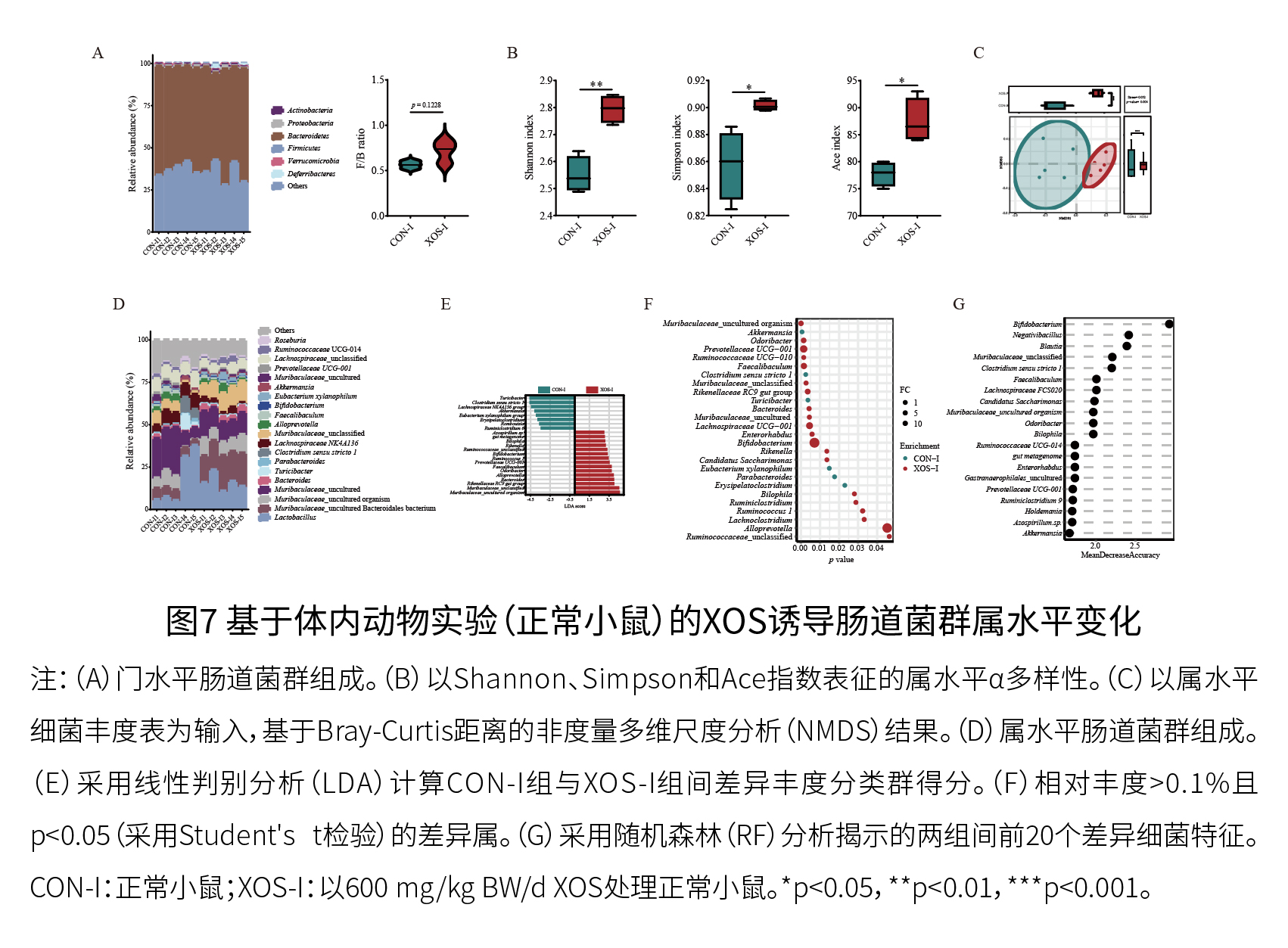
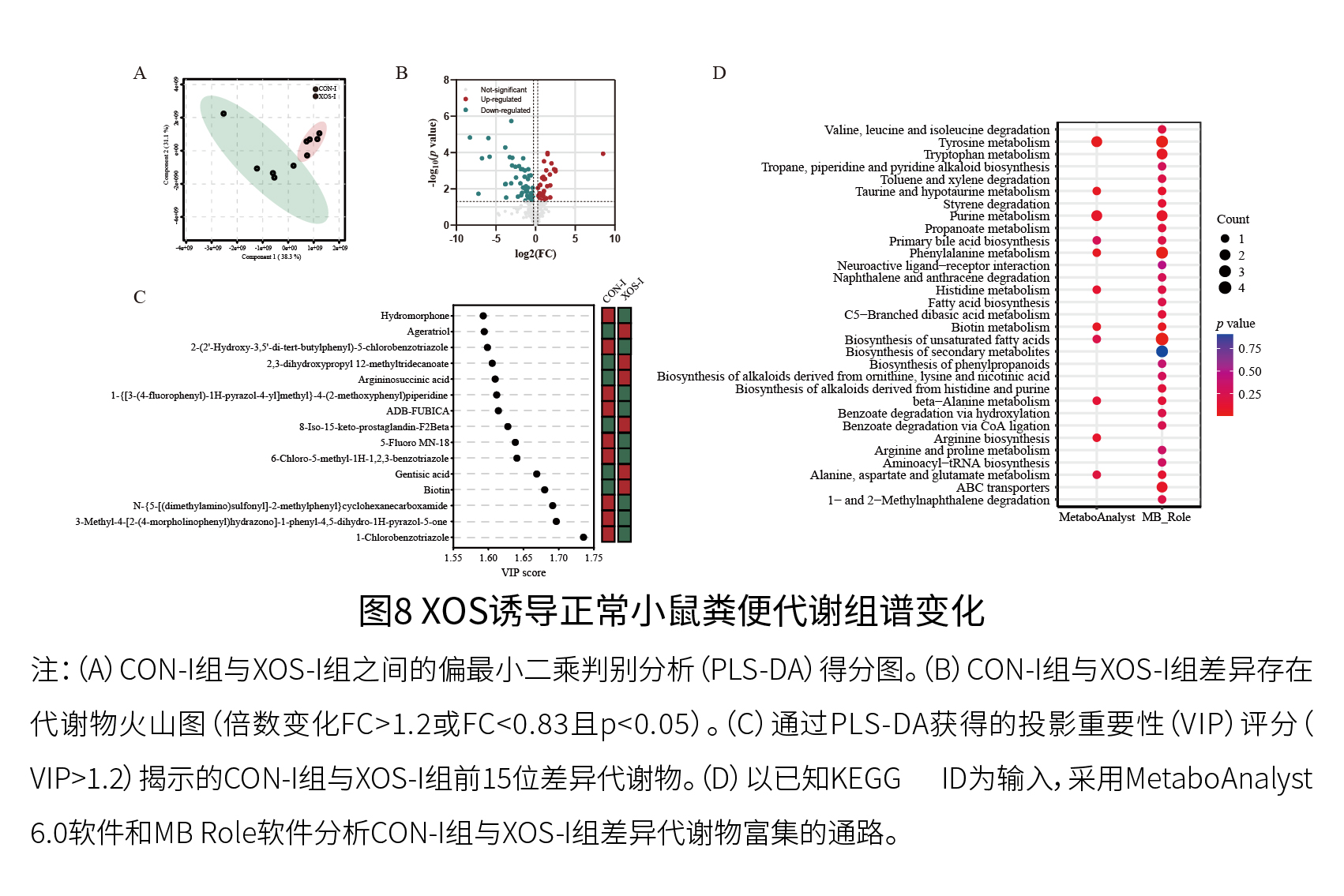
实验二:动物实验 (1)XOS有效缓解D-半乳糖诱导的衰老小鼠的衰老 如图7和8所示,与体外粪便发酵观察到的结果一致,XOS干预增加了关键XOS响应微生物属(双歧杆菌)的相对丰度,并影响了与宿主健康结果和/或衰老相关的代谢途径,包括精氨酸和脯氨酸代谢、精氨酸生物合成、色氨酸代谢以及与胆汁酸代谢相关的途径。总的来说,体外人类粪便发酵和体内动物实验(正常小鼠)的结果一致表明,XOS补充与粪便中与衰老相关的属、代谢物和代谢途径的变化密切相关。
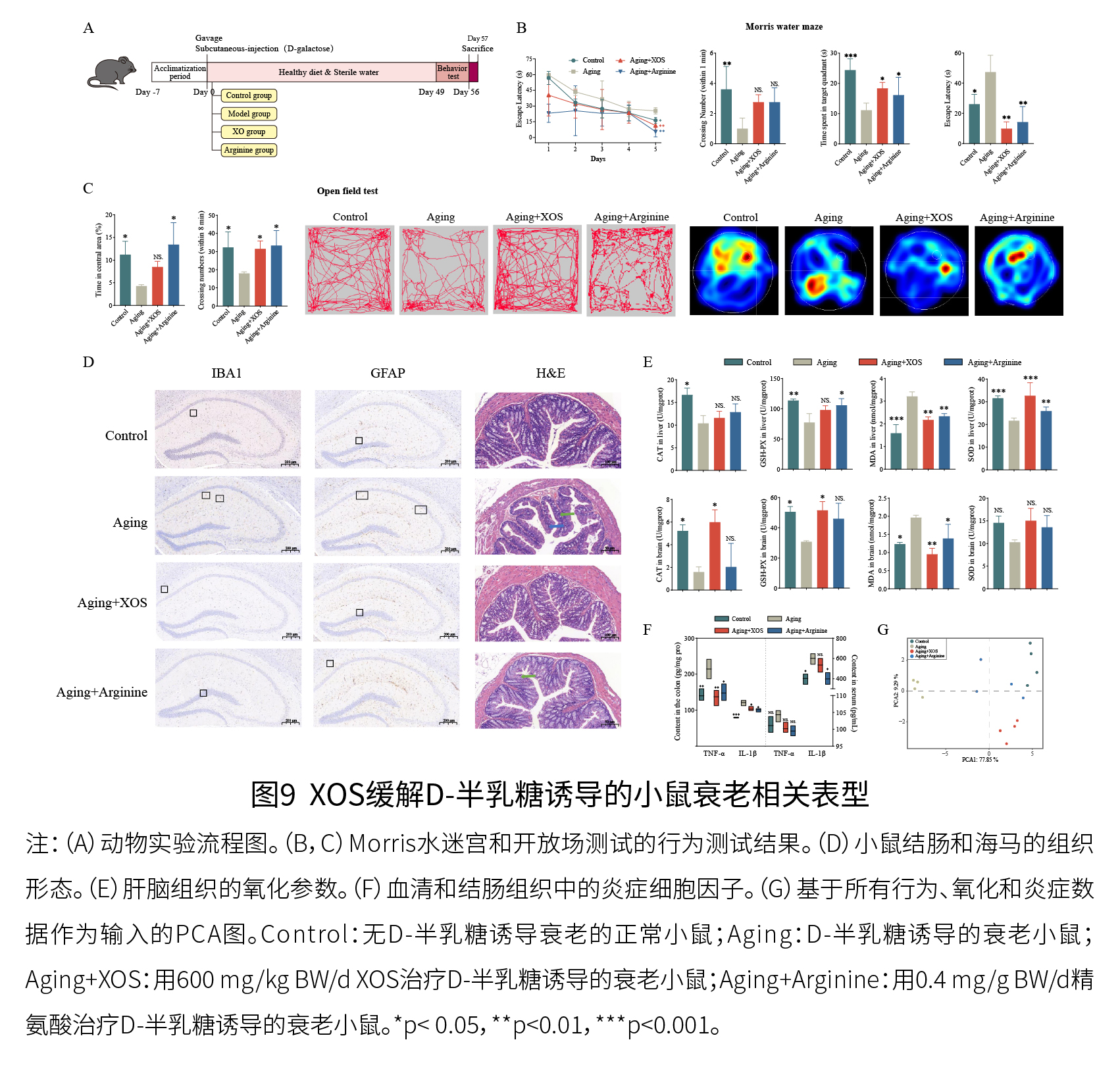
如图9A-C所示,与对照小鼠相比,D-半乳糖诱导的衰老小鼠表现出记忆力下降,而XOS干预缓解了在这一现象。如图9D所示,D-半乳糖诱导的衰老导致了衰老的小胶质细胞和星形胶质细胞的出现以及认知衰退,这些都可以通过XOS干预得到显著改善。另外,结肠H&E染色显示,D-半乳糖诱导的衰老可能导致结肠炎症,而XOS摄入可能减轻结肠的炎症状态。如图9E和9F所示,XOS补充显著恢复了脑组织中GSH-Px和CAT的抗氧化活性,并降低了MDA含量。在肝脏组织中,四个参数中有两个(SOD活性和MDA水平)在XOS干预后显著改善,XOS处理也显著降低了结肠中促炎细胞因子(TNF-α和IL-1β)的水平。
总之,XOS有效地缓解了D-半乳糖诱导的小鼠宿主衰老表型。体外粪便发酵实验、体内XOS补充(正常小鼠)以及体内衰老动物实验的功能验证一致表明,XOS对肠道微生物群和相关代谢的影响可以转化为改善宿主衰老表型。 试验结论 在本研究中,通过体外粪便发酵实验探讨了XOS对人体肠道微生物和代谢的影响。具体而言,探索了XOS转化在属和种水平上肠道微生物群的特征,以进一步阐明XOS的益生元作用,并鉴定了XOS响应的关键细菌(长双歧杆菌长亚种),以及XOS干预改变了有益细菌(扭链瘤胃球菌、双歧杆菌属和布劳特氏菌属)以及重要生理代谢物(有机酸、短链脂肪酸、色氨酸衍生物和胆汁酸代谢)的丰度,以及相关的代谢途径(精氨酸及脯氨酸代谢信号通路)。此外,研究不仅在分子水平上阐明了微生物对XOS反应的潜在机制,而且还表明肠道微生物群之间的代谢联系在XOS反应中起着重要作用,并确定了三组功能不同的肠道微生物群。鉴于XOS干预改变了与衰老相关的肠道微生物代谢物,进一步通过动物实验验证了XOS在缓解宿主衰老方面的作用。这些重要发现为基于体外粪便发酵的有益益生元筛选和XOS作为具有健康和衰老益处的安全有效的益生元提供了新的证据。 参考资料: Feng Y X, Xiao Y, Li X T, et al. Prebiotic roles and anti-aging effects of xylo-oligosaccharide: Keystone responsive bacteria and their metabolic interactions[J]. Food Research International, 2025, 215(9): 116673. |